

- 13.35 MB
- 2022-08-12 发布


- 1、本文档由用户上传,淘文库整理发布,可阅读全部内容。
- 2、本文档内容版权归属内容提供方,所产生的收益全部归内容提供方所有。如果您对本文有版权争议,请立即联系网站客服。
- 3、本文档由用户上传,本站不保证质量和数量令人满意,可能有诸多瑕疵,付费之前,请仔细阅读内容确认后进行付费下载。
- 网站客服QQ:403074932
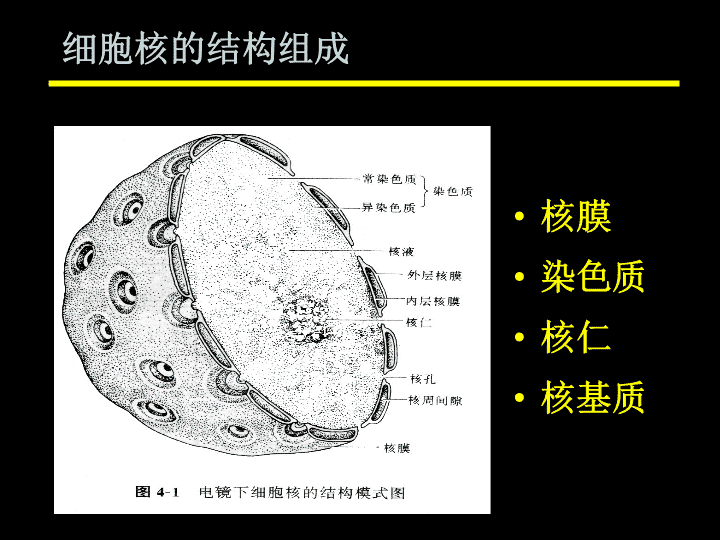
第八章细胞核(nucleus)\n细胞核是真核细胞内最大、最重要的细胞器,是细胞遗传(细胞内DNA复制的RNA转录的中心)与代谢的调控信息中心。细胞核的大小、位置和数量常因细胞的类型不同而有很大差异。核质比=细胞核的体积/细胞质的体积真核细胞通常只有一个细胞核,但肝细胞、软骨细胞、破骨细胞等可有几个到多个核。\n细胞核的结构组成核膜染色质核仁核基质\n四核仁一核膜和核孔复合体二核基质三染色质及染色体五 细胞核的功能\n第一节核膜核膜(nuclearmembrane)又称核被膜(nuclearenvelope),为双层膜结构。一、核膜的化学组成以蛋白质与脂质为主,含少量核酸。与内质网具相似成分。\n二、核膜的亚微结构不对称双层膜。核周间隙、核孔、核纤层。核周间隙异染色质核纤层核孔复合体细胞质外核膜内核膜核糖体膜整合蛋白\n外核膜(outernuclearmembrane)与粗面内质网相连,外表面附着核糖体。内核膜(innernuclearmembrane)无核糖体附着,内侧有一层致密的纤维蛋白网络,称为核纤层,有支持核膜的作用。(一)核膜的双层膜结构\n核周间隙指内外核膜之间的腔隙,宽约20-40nm,与内质网腔相通。(二)核周间隙(perinuclearspace)\n(三)核孔(nuclearpore)核孔复合体(nuclearporecomplex,NPC)\n胞质环(cytoplasmicring),外环核质环(nuclearring),内环,“核蓝”结构中央栓(centralplug)轮辐(spokes)结构模型—捕鱼笼式“核蓝”\n\n(四)核膜的主要功能1.区域化作用将细胞质与细胞核物质限定在特定区域,避免生命活动的彼此干扰(使RNA的转录及蛋白质的翻译在时间和空间上得以分开)。2.控制核质之间的物质转换与信息交流无机离子及小分子物质—以自由扩散,被动转运方式通过核孔复合体中央水性通道;DNA、RNA、酶类等大分子--由核孔复合体选择性主动转运。\n核转运受体与核定位信号核转运受体(nucleartransportreceptor):存在于核孔复合体上,介导选择性主动转运。核定位信号(nuclearlocalizationsignal,NLS):存在于被转运核蛋白上的一些短的氨基酸序列片段,为核转运受体识别的位点,引导蛋白质进入细胞核。主动转运过程中,核孔复合体上的酶水解ATP提供能量。\n\n核孔复合体上还存在识别RNA或RNA结合蛋白的受体,将转录产物RNA由细胞核转运到细胞质。核孔复合体的选择性转运具有双向性—核输入与核输出:细胞质细胞核DNA复制、RNA转录相关的酶类RNA、RNA结合蛋白等\n4.合成生物大分子外核膜上附着核糖体,参与蛋白质合成。3.在细胞分裂中参与染色体的定位与分离\n第二节核纤层与核骨架一、核纤层(nuclearlamina)紧贴内核膜的一层高电子密度纤维蛋白网,核内与核骨架相连,核外与中间纤维相连。\n核纤层由核纤层蛋白(lamin)构成。核纤层的作用:1.支持核膜,固定核孔位置;2.为染色质提供附着点;3.参与细胞分裂中染色质凝集的调节;4.与核膜的裂解和重建有关.\n二、核骨架(nuclearscaffold)又称核基质(nulcearmatrix),为间期核内除染色质、核膜、核仁外的由非组蛋白组成的纤维网架结构,形态类似细胞骨架。核骨架与核纤层以及细胞质中的中间纤维形成“核骨架-核纤层-中间纤维”体系,贯穿细胞核与细胞质之间。核骨架的主要化学成分:核基质蛋白和核基质结合蛋白两大类,200种以上。并含有少量RNA。\n核骨架的功能:1.与DNA复制有关。2.与基因表达有关。3.与RNA的转录后修饰及定向运输有关。4.与细胞分裂有关。5.与细胞分化有关。\n第三节染色质和染色体染色质(chromatin):指间期细胞核内由DNA、组蛋白、非组蛋白及少量RNA组成的线性复合结构,是间期细胞遗传物质存在的形式。染色体(chromosome):指细胞在有丝分裂或减数分裂过程中,由染色质盘绕压缩而成的棒状结构,也是分裂期遗传物质的存在形式。\n染色质与染色体是细胞核内同一种成分在细胞周期不同时相的不同表现形式。染色质:间期,不规则细线状,网状弥散于核内。染色体:分裂期,由染色质盘绕压缩而成的棒状结构。\n一、染色质和染色体的化学组成DNA组蛋白非组蛋白少量RNA为主(1:1)\n(一)DNA脱氧核糖核酸,双螺旋链,遗传信息的物质载体。每条染色体含一条线形的DNA分子。其序列分为:单一序列:单拷贝、不重复,占60%-65%。重复序列:多拷贝。中度重复序列:占20%-30%。高度重复序列:拷贝数大于105,占10%。多分布于着丝粒及端粒。\n3种特殊的DNA序列:复制起点序列:多个成串激活,同时复制,保持遗传连续性。着丝粒序列:连接姐妹染色单体,确保遗传稳定性。端粒序列:位于染色体端部,保持染色体独立及稳定。染色体DNA的三种功能元件,P185\n端粒复制起点着丝粒间期有丝分裂期间期复制眼动粒\n(二)组蛋白(histone)核小体组蛋白(nucleosomalhistone):H2A、H2B、H3和H4,帮助DNA卷曲形成核小体的稳定结构。H1组蛋白:H1,在构成核小体时H1起连接作用,赋予染色质以极性。\n真核生物染色体的基本结构蛋白,富含带正电荷的Arg和Lys等碱性氨基酸,属碱性蛋白质,可以和酸性的DNA紧密结合(非特异性结合);没有种属及组织特异性,在进化上十分保守。可以进行化学修饰:乙酰化、磷酸化和甲基化。在细胞周期的S期与DNA同时合成。组蛋白的特点:\n(三)非组蛋白(non-histone)染色体中除组蛋白外的其它蛋白。对DNA的识别具有特异性,从而调控基因的表达(与其磷酸化修饰关系密切)、促进染色质高级结构的形成等。种类多,有种属和组织特异性\n二、染色质的种类常染色质(euchromatin):间期核内处于分散状态,具有活跃DNA复制及转录功能的染色质组分,用碱性染料染色时着色浅。异染色质(heterochromatin):间期核内处于凝集状态,转录功能缺乏或不活跃的染色质组分,碱性染料染色时着色较深。\n异染色质的类型:结构异染色质(constitutive-heterochromatin)除复制期以外,在整个细胞周期均处于聚缩状态,形成多个染色中心。多定位于着丝粒、次缢痕。兼性异染色质(facultative-heterochromatin)在某些细胞类型或一定的发育阶段,原来的常染色质聚缩,并丧失基因转录活性,变为异染色质,如X染色体随机异染色质化失活。异染色质化可能是关闭基因活性的一种途径。为主\n异染色质常染色质\n常染色质异染色质间期染色程度浅深分布核中央,染色体臂核膜附近螺旋化程度低,疏松状态高,凝集状态DNA序列单一序列和中度重复序列(如组蛋白基因和tRNA基因)高度重复序列功能状态转录和翻译活跃转录和翻译不活跃常染色质与异染色质的比较\n活性染色质(activechromatin):具有转录活性的染色质,为常染色质。非活性染色质(inactivechromatin):没有转录活性的染色质,占大多数,包括常染色质与异染色质。染色质按功能状态不同分为:\n三、染色质的结构与装配染色质的基本结构单位—核小体(nucleosome)(一)染色质的一级结构--11nm染色质纤维由核小体串联组成,呈念珠状。\n核小体结构要点:每个核小体单位包括约200bp的DNA、一个组蛋白八聚体核心和一个H1组蛋白;八聚体由H2A、H2B、H3、H4各两分子形成,构成核小体的核心;DNA分子螺旋缠绕在核心颗粒表面,每圈83bp,共1.75圈,约146bp,两端被H1锁合;相邻核心颗粒之间为一段60bp的连接线DNA。\n\n核小体核心颗粒核心DNA连接部DNA连接部DNA连接部\n核小体与核小体连接形成串珠状的染色质纤维\n(二)染色质的二级结构--30nm染色质纤维\n螺线管(solenoid):由6个核小体螺旋围成一圈形成,H1组蛋白位于内部,调节其组装与稳定。螺线管侧面观螺线管顶面观\n(三)染色质的高级结构多级螺旋模型(multiplecoilingmodel)放射环模型(radialloopmodel)\n多级螺旋模型一级结构:核小体二级结构:螺线管三级结构:超螺线管(supersolenoid)四级结构:染色单体(chromatid)\nDNA核小体螺线管超螺线管染色单体压缩7倍压缩6倍压缩40倍压缩5倍\n非组蛋白构成染色体骨架(chromosomalscaffold),由骨架伸出的无数的DNA侧环;放射环模型30nm的染色质纤维折叠成环,沿染色体纵轴,由中央向四周伸出,构成放射环。\n袢环染色体支架\n四、染色体的结构分裂中期染色体形态及结构较稳定,可作为标准。\n染色单体(chromatid)中期染色体含两条染色单体,又称姐妹染色单体,以着丝粒相连,由间期复制后组装形成。\n2.着丝粒及动粒着丝粒(centromere):中期染色单体相互联系在一起的特殊部位,染色体主缢痕处,由高度重复异染色质组成。动粒(kinetochore):主缢痕两侧的特化圆盘状结构,与纺锤体微管相连,由蛋白构成。\n着丝粒--动粒复合体包含3个结构域动粒结构域(kinetochoredomain)中央结构域(centraldomain)配对结构域(pairingdomain)内板(innerplate)中间间隙(middlespace)外板(outerplate)纤维冠(fibrouscorona)\n动粒结构域染色体臂着丝粒(主缢痕)中央结构域配对结构域纤维冠外板中间间隙内板\n位于有些染色体短臂末端的球形或棒形结构,通过次缢痕与短臂相连,异染色质组成,稳定。3.次缢痕染色体上除主缢痕外的缢缩部位。4.随体\n5.端粒(telomere)染色体末端的特化部位,由高度重复的短序列组成,高度保守。\n维持染色体的稳定性,防止染色体末端粘连。起细胞分裂计时器的作用。染色体每复制一次,端粒核苷酸减少50~100bp,使端粒长度逐渐减少,导致衰老发生。端粒的作用:多数肿瘤细胞中存在端粒酶活性,可不断复制合成端粒,从而无限增殖而不衰老。正常细胞中无端粒酶活性。\n染色体各部分的名称随体次缢痕主缢痕端粒长臂短臂\n核型:指染色体组在有丝分裂中期的表型,包括染色体数目、大小、形态特征的总和。染色体显带技术:染色体沿纵轴方向显示宽窄深浅不一的条带。根据染料及显色方法不同,有Q带、G带、R带和C带等。五、核型与带型\n\nABCDEFG\n\n\n\n第四节核仁核仁(nucleolus)细胞核中均质、无包膜的球形小体,一个或多个,多位于核的一侧。核仁是细胞中形成核糖体的结构。\n一、核仁的化学组成蛋白质RNADNA微量的脂类占80%,包括核糖体蛋白、染色质组蛋白和非组蛋白、酶类。\n二、核仁的结构核仁相随染色质纤维结构颗粒成分核仁基质\n1.核仁相随染色质核仁周围染色质:异染色质。核仁内染色质:常染色质,占主要部分。含rDNA,为核仁组织者。\n核仁组织者(区)Nuclearorganizer核仁内染色质上含有成串的rRNA基因,这个染色体区段称为核仁组织者。它们在间期时相互融合形成核仁,构成的区域称为核仁组织者区(NOR);在分裂中期时存在于特定染色体的次缢痕处。人类的rRNA基因位于13、14、15、21、22号5对染色体的端部。\nNOR有丝分裂时核仁为什么会消失?染色质\n也称核仁骨架,与核骨架相通。2.纤维结构由致密的纤维构成,主要成分是正在转录的rRNA,以及核糖体蛋白和RNA结合蛋白等。3.颗粒成分直径15-20nm的颗粒,是rRNA进一步加工成熟的区域,是不同加工阶段核糖体亚单位前体。4.核仁基质\n三、核仁周期核仁的动态变化:核仁随细胞周期的进行而呈现周期性变化(形成和消失)。染色体含rDNA的核仁组织区解旋和伸展,重新开始形成核仁。间期分裂期染色质浓缩,核仁组织者区缩回染色体,纤维成分和颗粒成分分散在核质中,整个核仁缩小继而消失。\n四、核仁的功能合成rRNA装配核糖体\n1.rRNA的合成5SrRNA5.8SrRNA18SrRNA28SrRNA在核仁外染色体上合成在核仁内合成位于NOR的rDNA是rRNA的信息来源。形态特征:“圣诞树”样结构。\n编码3种rRNA的基因紧密连锁为一个转录单位多个转录单位串联为基因簇,中间以一段不转录DNA相隔\n转录时,RNA聚合酶沿DNA链排列,转录好的rRNA分子从聚合酶处伸出,沿转录方向愈近末端愈长,呈“圣诞树”状。\nrRNA前体的加工rRNA在纤维结构处合成,它与蛋白质形成RNP复合体(核糖核蛋白复合体),在颗粒结构处加工。45SrRNA前体20S+32S18S28S5.8S+\n2.核糖体的组装18SrRNA40S核糖体小亚基5SrRNA28S+5.8SrRNA60S核糖体大亚基\n核糖体的成熟作用只发生在转移到细胞质以后,从而阻止有功能的核糖体与核内加工不完全的hnRNA分子接近。\n第五节细胞核的功能主要功能:遗传信息的贮存、复制、传递与表达。DNA复制RNA转录\n一、遗传信息的贮存什么是遗传信息?——DNA分子中碱基(核苷酸)的排列顺序。基因:DNA分子中具有一定生物功能的区段,是携带遗传信息的结构单位,控制某一特定的性状。编码:mRNA—合成蛋白质tRNA、rRNA—协助合成蛋白质\n遗传信息的流动:DNA转录翻译RNA蛋白质\n复制复制中心法则CentraldogmaDNARNA蛋白质翻译转录逆转录\n二、遗传信息的复制DNA复制(replication):DNA分子在DNA合成酶系的作用下,合成与自身分子结构相同的子代DNA的过程。复制亲代DNA子代DNA\n(一)DNA复制所需的酶及蛋白质DNA解旋酶:利用ATP供能,作用于氢键,解开DNA双螺旋链。DNA单链结合蛋白:在复制中维持模板处于单链状态并保护单链的完整。引物酶:复制起始时催化生成RNA引物的酶。\nDNA聚合酶(DNApolymerase,DNA-pol):原核细胞–DNA聚合酶I、II、III真核细胞–DNA聚合酶α、β、δ、ε。活性:1.53聚合活性2.53核酸外切酶活性3.35核酸外切酶活性(DNA聚合酶I&DNA聚合酶δ)\n53DNA聚合活性\n5´AGCTTCAGGATA3´|||||||||||3´TCGAAGTCCTAGCGAC5´35外切酶活性53外切酶活性?切除RNA引物、突变的DNA片段等。辨认错配的碱基对,并将其水解。核酸外切酶活性\n连接酶:催化相邻DNA片段间的3’-OH与5’-PO4间形成磷酸二酯键,从而使DNA片段连接在一起。DNA连接酶ATPADPHO5’3’5’3’3’5’5’3’\n端粒酶:在DNA复制终止时发挥作用的一种酶,由RNA与具有反转录酶活性的蛋白质组成。功能:以自带RNA片段为模板,催化补齐DNA端粒末端的重复序列,避免DNA在复制中逐渐缩短。普通的正常细胞中无端粒酶活性。\n(二)DNA复制的特性1、半保留性复制形成的子代DNA中,一条单链来自亲代模板DNA,另一条单链则为完全重新合成的互补链。这种复制方式称为半保留复制。\nAGGTACTGCCACTGGTCCATGACGGTGACCCCACTGGGGTGACCAGGTACTGTCCATGACTCCATGACAGGTACTGAGGTACTGCCACTGGTCCATGACGGTGACCAGGTACTGCCACTGGTCCATGACGGTGACC+母链DNA复制过程中形成的复制叉子代DNA\n3’5’引物酶两条DNA单链的复制从固定起点开始,向两侧相反方向推进(各自分别沿新合成链的5’3’方向)。2、双向性复制叉(replicationfork)\n3、多起点性真核生物DNA复制有多个起始点。习惯上把具有一个DNA复制起点的复制单位称为复制子(replicon)。因此真核生物的DNA复制是多复制子的双向复制,最终互相连接。\n\n4、不连续性DNA聚合酶催化DNA链合成方向:5’3’5’5’3’复制子3’\n3535解链方向3´5´3´3´5´前导链(leadingstrand)后随链(laggingstrand)RNA引物\n前导链(leadingstrand):合成方向同复制叉推进方向一致,连续合成。后随链(laggingstrand):合成方向同复制叉推进方向相反,短片段、不连续合成。复制中的不连续片段称为岡崎片段(okazakifragment)。\n\n5、不同步性不同复制子在复制时间上存在差异。常染色质复制早于异染色质。\n复制过程简图引物酶前导链\n三、遗传信息的转录转录(transcription):以DNA为模板合成RNA的过程。转录RNADNA\nDNA双链中作为转录模板的一股单链称为模板链(templatestrand),也称作反义链(antisensestrand)。相对的另一股互补单链,其碱基序列与转录产物相同,称为编码链(codingstrand),也称为有义链。5335模板链编码链编码链模板链转录方向转录方向\n复制和转录的区别A-U,T-A,G-CA-T,G-C配对mRNA,tRNA,rRNA子代双链DNA(半保留复制)产物RNA聚合酶(RNA-pol)DNA聚合酶酶NTPdNTP原料模板链转录(不对称转录)两股链均复制模板转录复制A-U,T-A,G-CA-T,G-C配对mRNA,tRNA,rRNA子代双链DNA(半保留复制)产物RNA聚合酶(RNA-pol)DNA聚合酶酶NTPdNTP原料模板链转录(不对称转录)两股链均复制模板转录复制\n(一)转录的基本过程—起始、延长、终止1.转录的起始原核生物的RNA聚合酶:4种亚基α,β,β’,。其中亚基识别转录起始点,称为转录的起始因子。核心酶全酶\n真核生物的RNA聚合酶(3种):RNA聚合酶I、II、III.亚基种类多,结构复杂。种类IIIIII分布核仁核基质核基质产物45SrRNAhnRNA5SrRNA、tRNA前体对鹅膏蕈碱的反应不敏感极敏感中度敏感\n启动子(promoter):RNA聚合酶开始结合于模板DNA的部位。注意区分:启动子是模板DNA上最开始与RNA聚合酶结合的一段DNA序列;转录的起始因子是RNA聚合酶上的一个蛋白质亚基。\n原核细胞中RNA聚合酶结合启动子:TATAATGATATTAC启动子识别部位启动子结合部位(Pribnowbox)转录起始因子亚基识别启动子识别部位,并介导RNA聚合酶全酶结合于启动子结合部位,从而启动转录。\n真核细胞中RNA聚合酶结合的启动子:3种RNA聚合酶分别识别各自的启动子序列。RNA聚合酶II的启动子TATA框CAAT框------CAAT-----TATA转录起始点-26~30bp-75bp结构基因RNA聚合酶结合部位,控制转录起始的精确性。与RNA聚合酶结合,控制转录起始的频率。\n转录因子(transcriptionfactor,TF):转录过程中与DNA特殊序列结合从而调控转录进程的蛋白质。在真核细胞中,只有当启动子的DNA序列上结合一个或多个特异的DNA结合蛋白时,才成为有功能的启动子,可被聚合酶所识别。RNA聚合酶—转录因子—启动子真核细胞中RNA聚合酶与启动子的结合需要转录因子参与。\n转录因子:TFIIBTFIIDTFIIETFIIS-RNApol-TATA框形成复合物,启动转录。促进转录延伸。\n2.转录的延伸原核细胞中:转录起始后,亚基从全酶上脱落,核心酶沿模板链移动,RNA链沿5’3’方向延长。真核细胞中:转录延长过程与原核细胞大致相似,转录因子TFIIE与TFIIS有促进延长的作用。\n3.转录的终止RNA聚合酶在DNA模板上移动到终止位点,停止作用,产物RNA链从模板上脱落。依赖ρ因子的转录终止非依赖ρ因子的转录终止原核细胞中的转录终止\n依赖ρ因子的转录终止ρ因子识别DNA上终止序列并与之结合当RANpol移至终止位点,与ρ因子结合RAN链脱离模板,转录终止\n非依赖ρ因子的转录终止DNA模板上靠近终止处,有些特殊的碱基序列,转录出RNA后,RNA产物局部形成发夹结构,引起转录终止。5´pppG5335RNA-pol\n(二)转录后的加工DNA分子上转录出RNA的区段,称为结构基因(structuralgene)。真核细胞的结构基因一般是不连续的,称为断裂基因(splitgene),由编码蛋白质的序列和非编码蛋白质的序列构成。CABD编码区A、B、C、D非编码区\n外显子(exon)和内含子(intron)外显子断裂基因上可编码蛋白质,表达为成熟RNA的核酸序列。内含子在断裂基因中出现在外显子之间,在剪接过程中被除去的核酸序列,不参与编码蛋白质。\n1.原核细胞中转录后的加工mRNA:不需剪切加工。tRNA:切除5’及3’端点附加序列,在3’端附加CCA序列,对特定碱基进行化学修饰。rRNA:将rRNA前体剪切成5S、16S、23SrRNA。原核细胞的结构基因是连续的,不含内含子。\n(1)mRNA前体的加工2.真核细胞中转录后的加工核不均一RNA(heterogeneousnuclearRNA,hnRNA)成熟的mRNA戴帽剪接加尾\n戴帽在hnRNA5’端加上帽子结构,即7-甲基鸟嘌呤核苷-5’-三磷酸鸟苷(m7GpppG……)。在hnRNA合成到30个核苷酸时开始形成。作用:封闭hnRNA5’端,稳定hnRNA,防其被降解;为核糖体小亚基识别位点,帮助mRNA与核糖体小亚基结合。\n鸟苷酸转移酶甲基化酶\n加尾在3端加上含100~200个A的多聚腺苷酸尾巴(polyAtail)。作用:稳定mRNA,并帮助其由核输出至胞质。AAUAAA\nAAUAAA\n剪接切除hnRNA中的内含子,拼接外显子。剪接信号剪接体互相识别保证精确剪接内含子5’端的GC,3’端的AG——剪切位点内含子3’端上游UACUAAC序列中的A——分支点、交叉点\n小分子RNA:U1-U6(smallnuclearRNA,snRNA)核内的蛋白质小分子核糖核蛋白颗粒(smallnuclearribonucleoproteinparticle,snRNP)剪接体:U1snRNP、U2snRNP……U6snRNP\n剪接过程①U1snRNP结合5’剪切位点;U2snRNP结合分支点。②U4、U5、U6snRNP以复合体形式结合于内含子,形成剪接体。\n③分支点上的A向5’端剪接点靠近。④内含子与外显子在5’端剪接点处断开,断开的内含子5’端与A相连,形成套索状结构。\n⑤5’端外显子的3’-OH与3’端外显子的5’-PO4结合,内含子3’剪切点被切除。⑥剪切体及套索结构脱离,剪接过程结束。剪切过程中需要ATP提供能量。\nhnRNA的选择性剪接234561234123561\n(2)rRNA前体的加工转录(RNApolI)45SrRNA前体18SrRNA28SrRNArDNA内含子内含子28S5.8S18S剪接5.8SrRNA5SrRNA\n(3)tRNA前体的加工tRNA前体RNApolⅢDNA\nRNaseP核酸内切酶前导序列内含子tRNA核苷酸转移酶转运Aa的结合位点\n其它碱基修饰\n(三)转录的调节真核细胞中转录调节的主要方式----基因附近的DNA序列与相关的DNA结合蛋白结合,促进或抑制靶基因的转录。顺式作用元件(cis-actingelement):对基因转录有调节活性的DNA序列。反式作用因子(trans-actingfactor):与顺式作用元件结合的蛋白质,又称转录因子。\n顺式作用元件启动子:包括TATA框、CAAT框及GC框(GGGCGG)等;增强子:能增强启动子功能活性、加速基因转录的DNA序列,位置比较自由(可位于基因上下游、或内含子中,距离可远可近)。由多个增强体组成。沉默子:与增强子作用相反的负性调控元件,阻遏基因转录。\n转录起始点TATA盒CAAT盒GC盒增强子顺式作用元件(cis-actingelement)转录终止点外显子内含子启动子沉默子\n反式作用因子通用(基本)转录因子:结合启动子TATA框,帮助RNApol与启动子结合,参与转录启动。在3种RNApol间通用。特定转录因子:与特定顺式作用元件结合,调节特定基因的转录。转录激活因子:多为增强子结合蛋白;转录抑制因子:多为沉默子结合蛋白,或通过结合激活因子等抑制基因的表达。\n反式作用因子在结构上一般具有DNA结合域、转录激活域、其他蛋白结合域三个功能域;DNA结合域以非组蛋白形成不同结构模式:锌指结构(Zincfinger)α螺旋-转角-α螺旋结构(helix-turn-helixmotif)亮氨酸拉链基序(Lencinezippermotif,ZIP)螺旋-环-螺旋结构(helix-loop-helixmotif,HLH)\n螺旋-环-螺旋亮氨酸拉链螺旋-转角-螺旋锌指\n思考题(一)1、解释并区别下列名词:着丝粒与动粒主缢痕与次缢痕核小体与核心颗粒端粒与随体染色质与染色体核仁与核仁组织区核骨架与细胞骨架外显子与内含子转录因子与启动子与转录的起始因子顺式作用元件与反式作用因子断裂基因增强子\n2、简述核膜的结构和功能。3、简述染色质的化学组成。4、简述染色体的两种结构模型。5、常染色质与异染色质在结构和功能上有何异同?6、简述核仁的超微结构和功能。7、复制和转录有何区别?8、原核生物的启动子与真核生物RNA-polⅡ的启动子有何区别?9、真核生物mRNA的转录后加工包括哪些内容?思考题(二)\nTheEnd