

- 8.66 MB
- 2022-08-12 发布


- 1、本文档由用户上传,淘文库整理发布,可阅读全部内容。
- 2、本文档内容版权归属内容提供方,所产生的收益全部归内容提供方所有。如果您对本文有版权争议,请立即联系网站客服。
- 3、本文档由用户上传,本站不保证质量和数量令人满意,可能有诸多瑕疵,付费之前,请仔细阅读内容确认后进行付费下载。
- 网站客服QQ:403074932
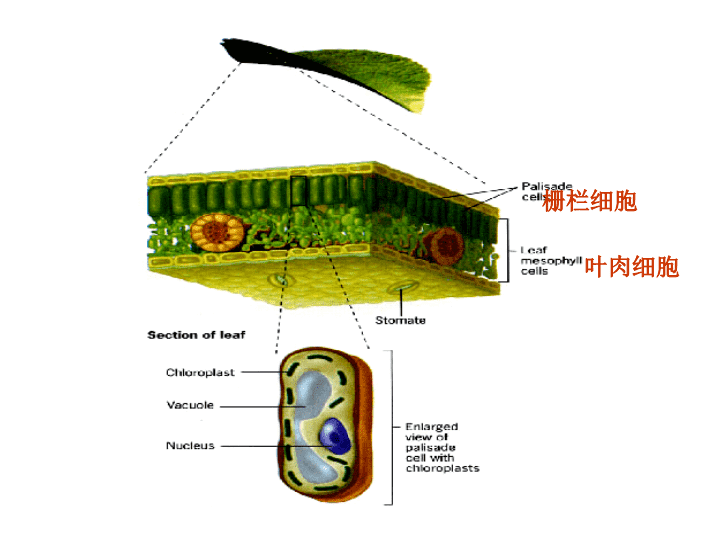
第二节叶绿体\n一叶绿体的形态、大小和数目形状:多呈香蕉形;直径:3~6μm,厚:2~3μm。数目:随种类和环境不同而差异巨大。\n栅栏细胞叶肉细胞\n二叶绿体的结构叶绿体膜/被膜(chloroplastmembrane/envelope)类囊体(thylakoid)基质(stroma)\n1叶绿体外被(外膜,膜间隙和内膜)外膜:高透性,核苷、磷酸衍生物可透过;内膜:透性很低,选择性强,是细胞质和叶绿体基质间的功能屏障;含转运体(translocator)以转运代谢物。膜间隙:厚约10~20nm,与内、外膜一起形成叶绿体外被。\n2类囊体类囊体:叶绿体基质中由单位膜封闭成的扁平小囊,一般沿叶绿体长轴平行排列;基粒(grana):由类囊体叠垛而成;基粒类囊体:基粒中的类囊体;基质类囊体:基粒间的类囊体.基粒类囊体\n类囊体膜上的主要组分①捕光色素;②PSⅡ(photosystemⅡ)复合物:由核心复合物和捕光复合物组成;③PSI(photosystemI)复合物;④细胞色素bf复合物(cytochromebfcomplex);⑤CFo-CF1ATP酶:CFo:H+通道;CF1:酶活性部位,需二硫苏糖醇和Mg2+激活。\n叶绿素的结构及其吸收光谱卟啉环\n\n\n3叶绿体基质基质:叶绿体内膜和类囊体之间的无定形物。主要成分:可溶性蛋白质(多为代谢酶,以RuBPase最多);核糖体、RNA和DNA;淀粉粒、脂滴及植物铁蛋白(储存Fe)。\n三叶绿体的主要功能—光合作用光合作用(photosynthesis):绿色植物叶肉细胞的叶绿体吸收光能,把CO2和水转化成有机物,并释放出氧的过程。光反应暗反应光合作用原初反应电子传递和光合磷酸化碳同化\n1原初反应(primaryreaction)原初反应:叶绿素分子被光激发至引起第一个光化学反应为止的过程。包括光能的吸收、传递和转换,完成光能到电能的转变。捕光色素/天线色素:大多数Chla和Chlb、Car.、Xanthophyll,只吸收光,不能发生光化学反应;反应中心色素:处于特殊状态的Chla,将光能转变为化学能,(P700(PSI),P680(PSⅡ));反应中心:进行光合作用的最小单位,包括反应中心色素、原初电子供体(D)和原初电子受体(A)。\n光反应中心和天线色素\n捕光色素捕获光能反应中心色素(激活)共振原初反应的过程:产生高能电子电荷分离\n电子供体电子受体\n2电子传递和光合磷酸化\n电子传递链的组成①PSⅡ(photosystemⅡ)复合物:由核心复合物(D1,D2,P680)、捕光复合物和放氧复合体组成;光解水,还原PQ②PQ:电子载体和H+载体,将H+泵入类囊体腔③Cytb-f复合物:④PSI(photosystemI)复合物;⑤Fd\n非循环式电子流(Z型):P680P700NADP+循环式电子流:P700FdCytb6P7001)电子传递的两条通路NADP+的浓度不足时均可产生跨类囊体膜的质子浓度差,与磷酸化偶联形成ATP;\n2)光合磷酸化光合磷酸化:由光照所引起的电子传递和磷酸化作用相偶联生成ATP的过程。将电能转化为活跃的化学能;类型:非循环式光合磷酸化:产物为ATP、NADPH和O2;循环式光合磷酸化:产物仅为ATP。\n光合磷酸化的作用机理(化学渗透学说):一对电子从P680经P700传至NADP+,在类囊体腔中增加4个H+(2个源于H2O光解,2个由PQ从基质转移而来),在基质中一个H+又被用于还原NADP+,所以类囊体腔内有较高的H+(pH≈5,基质pH≈8),形成质子动力势。H+经ATP合酶,渗入基质、推动ADP和Pi结合形成ATP。\n叶绿体和线粒体中ATP合成的异同同:ATP合酶的结构相似,具F1头部和F0基部;ATP形成都由H+流推动;光合和氧化磷酸化中都需要完整膜。不同:叶绿体,1对电子2次穿膜,在基质中摄取3H+(2个过膜,1个被NADP+吸收)在类囊体腔中产生相当于5H+(加上水光解产生的2个),每3H+生成1个ATP;线粒体:1对电子3次跨膜,将基质中的8H+抽提到膜间隙中,每2H+穿过F1-F0ATP酶,生成1个ATP分子。\n绿叶能发电返回美国、日本的农业科学家实验发现,绿叶可以发电。他们是利用绿叶在早晨八九点钟的太阳光照射下的光电效应,测出电流的;如果绿叶受光面积增大,则绿叶所发的电流也可增强。绿叶中的叶绿体发电的优点在于无论春夏秋冬、晴阴雨雪,它都能吸收光并转化为电能,因此专家认为,如果在房屋周围广泛植树,在墙边种植青萝紫藤、爬山虎等植物构成厚厚的多层覆盖的叠式植物群,不仅房舍周围绿树成荫,环境大为改善,而且每户人家还可自成体系地建成一个绿叶发电厂,从而为人类提供新的绿色能源\nPhotosynthesisrobot\n\n3光合碳同化利用光反应产生的ATP和NADPH(存在于叶绿体基质中),使CO2还原为糖类等有机物,将活跃的化学能转换为稳定的化学能,存于有机物中。此过程不直接需光,也叫暗反应,在叶绿体基质中进行。种类:◆卡尔文循环(Calvincycle)(C3途径);◆C4途径或Hatch-Slack循环;◆景天科酸代谢途径:(干旱地区的景天科植物)\n卡尔文循环羧化还原RuBP的再生\n光合作用与呼吸作用比较\n线粒体和叶绿体的比较叶绿体内膜并不向内折叠成嵴;内膜不含电子传递链;除了膜间隙、基质外,还有类囊体;捕光系统、电子传递链和ATP合成酶都位于类囊体膜上。\n四叶绿体DNA叶绿体DNA(ctDNA)的基本性质和功能性质:高等植物叶绿体DNA(chloroplastDNA,cp或ctDNA)一般是双链环状分子,其周长随种类而不同,大约在40~60µm,含200~2500kbp(千碱基对)。功能:编码叶绿体rRNA、tRNA和一些核糖体蛋白的基因;编码PSⅡ、PSI、Cytbf复合体、CF1—CF0ATP合酶和Rubisco等的一些亚单位。\n2参加叶绿体组成的蛋白质来源:由ctDNA编码,在叶绿体核糖体上合成;由核DNA编码,在细胞质核糖体上合成;由核DNA编码,在叶绿体核糖体上合成。\n叶绿体蛋白的运送与装配:定位于基质的蛋白:由转运肽(transitpeptides)牵引进入叶绿体;定位于类囊体蛋白:含两个信号序列;定位于类囊体膜的蛋白:叶绿体自身合成的蛋白。\n五叶绿体的发育和增殖叶绿体由前质体(proplastid)分化而来。增殖:分裂增殖,由中部向内收缩而分裂;幼龄叶绿体可分裂;但成熟叶绿体不能分裂。\n思考题1.试述线粒体和叶绿体的结构特征并比较其异同点。2.试述线粒体的氧化磷酸化和叶绿体的光合磷酸化作用。3.为何说ATP合酶是氧化磷酸化的关键装置?4.氧化磷酸化偶联机制的化学渗透假说的主要论点是什么?有哪些依据?5.为何说线粒体和叶绿体是半自主性细胞器?6.为何说在细胞质中线粒体蛋白质合成后输送至线粒体各部位是一个多步骤、由多种蛋白参与且需能量的过程?\n试从细胞或亚细胞水平举一例说明高等真核生物是如何实现结构和功能的相互统一的?