

- 6.37 MB
- 2022-08-12 发布


- 1、本文档由用户上传,淘文库整理发布,可阅读全部内容。
- 2、本文档内容版权归属内容提供方,所产生的收益全部归内容提供方所有。如果您对本文有版权争议,请立即联系网站客服。
- 3、本文档由用户上传,本站不保证质量和数量令人满意,可能有诸多瑕疵,付费之前,请仔细阅读内容确认后进行付费下载。
- 网站客服QQ:403074932
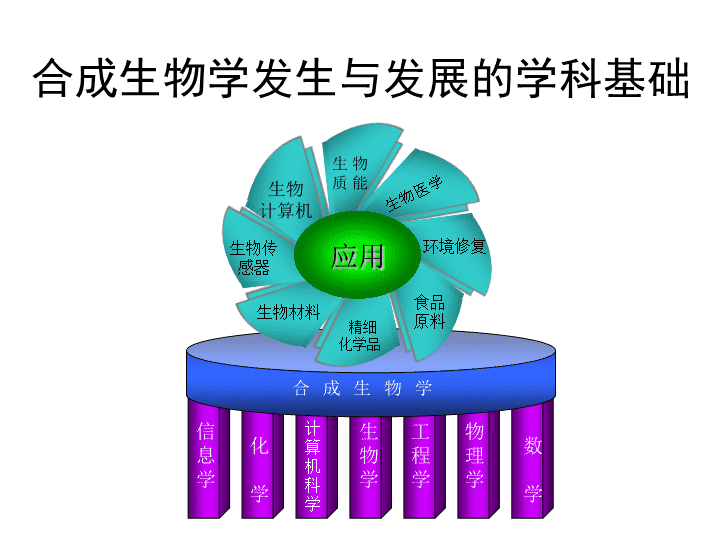
●维基百科全书:合成生物学旨在设计和构建工程化的生物系统,使其能够处理信息、操作化合物、制造材料、生产能源、提供食物、保持和增强人类的健康和改善我们的环境。http://en.wikipedia.org/Synthetic_biology●简单地说,合成生物学是通过设计和构建自然界中不存在的人工生物系统来解决能源、材料、健康和环保等问题。合成生物学的定义1新的生物零件、组件和系统的设计与建造;2对现有的、天然的生物系统的重新设计\n基因、基因组和基因组学:基因(Gene):遗传功能的单位,是编码蛋白质或RNA分子的一段DNA序列.基因组(Genome,来自Gene+chromosome)所有DNA分子的总和(分子遗传学定义)基因组学(Genomics):研究基因组结构与功能的科学\n合成生物学发生与发展的学科基础数学物理学工程学生物学计算机科学化学信息学合成生物学生物质能生物医学环境修复精细化学品食品原料生物材料生物传感器生物计算机应用\n合成生物学的研究内容生物大分子的合成与模块化生物基因组的合成、简化与重构合成代谢网络遗传/基因线路的设计与构建细胞群体系统及多细胞系统研究数学模拟和功能预测123465\n合成生物学的意义加速合成生物系统工程化的进程需要工程化、标准化的策略,将研究人员从日复一日的重复性操作中解脱出来。验证和深化对于生物现象的理解“合成”将是“分析”的必要补充。成功固然会帮助我们建立合成生物学的基本原则和生物系统的工程化技术;失败也是人类的理解与自然生物本质间存在鸿沟的直接佐证,并会为我们如何更好的理解和运用源自于自然的技术提供指引。\n合成生物学区别于现有生物学其他学科的主要特点是“工程化”2004年ScientificAmerican杂志发表了题为“合成生命”一文,以图文并茂的形式阐述了合成生物学的内涵及工程性质2005年MIT的Endy发表了“工程生物学的基础”的综述论文,明确提出工程中常用的“标准化”、“复杂系统解耦”、“概念抽象化”做法,并清楚地将合成生物学涉及的生物系统分成DNA、零件、装置、系统这样4个层次2006年ScientificAmerican以“THEBIOFABGROUP”(生物制造研究组,包括合成生物学领域的9位领军人物)为作者,发表了“工程生命:组建生物制造”的长篇分析评论性文章,在正文开始之前用大字突出强调:从有关工程领域的成功经验中所学到的原理及实践可帮助将生物技术从专门的工艺转化为一个成熟的工业。\n美国普林斯顿大学电子工程系与分子生物学系的Weiss等发表了题为“合成生物学:对于一个刚出现学科的新的工程作用”的综述。概括了合成生物学新学科的基本性能以及与其它工程学科相比的独特性;讨论了从生物装置、模块、细胞到多细胞系统各个层次进行设计和建造工程细胞的方法Panke等发表了“合成生物学-将工程置入生物学”的述评,归纳出合成生物学的研究框架,特别突出了系统设计与系统加工制造的重要性。对于作为合成生物学支撑技术的加工制造,包括标准化克隆、从头DNA合成、提供工程底盘机架(chassis)等,进行了详细讨论。为了减少生物系统的复杂性,一个选择是减小宿主(这里比作chassis)的基因组———即构建出“最小基因组”,以便用于合成生物学研究\n美国杜克大学生物医学工程系You的研究组及美国哈佛医学院系统生物学系Silver的研究组最近分别发表了题为“经由设计的生物学:细胞组分及特性的还原与合成”及“设计生物系统”的综述,突出体现了“设计”在合成生物学中的重要作用。Barrett等人评述了系统生物学与合成生物学的关系,强调指出系统生物学是基因组尺度合成生物学的基础。\n从化学工程和电子工程得到的借鉴通常人们认为DNA遗传密码是指挥控制生命的软件(software),而细胞膜及细胞内所有的生物机器被认为是生命的硬件(hardware,在合成生物学中也常称为wetware)。这种对生命系统软件与硬件的认识可以借助于电子工程(大规模集成电路技术、电子计算机硬件与软件技术等)的研究方法、基本技术与工具。就像技术人员现在用标准化的、现成的电子元件组装成计算机一样,合成生物学工作者预计有一天,工程师可以将充分表征的生物原件组装成健壮的宿主生物体,其具有特定的生物功能。\n美国国家自然科学基金资助的合成生物学工程研究中心(SynBERC)为此确定出4个研究开发方面:零件(parts)、装置(devices)、底盘机架(chassis)和人工实践(humanpractices)\n在建造计算机时,重要的是将全部模块装置(例如:母板、盘驱动器、输入输出部件等)集成到底盘机架内。对于合成生物学来说,创建一个细胞底盘机架(cellularchassis)用来安装我们设计的生物零部件是非常重要的。在细胞底盘机架中可以集成来自各个子系统的信号以使其有复杂的细胞功能。细胞底盘机架必须为细胞生长及组件工作提供各种组分,应该有各种标准的连接,而且足够稳定以便能在工业上应用。大肠杆菌可以认为是一个性能优良的微生物,可用作一个细胞底盘机架。\n工程化研究策略自上至下(逆向工程)和自下至上(前/正向工程)。自上至下策略主要用于分析阶段,试图利用抽提和解耦方法降低自然生物系统的复杂性,将其层层凝练成工程化的标准模块。自下至上的策略是指通过工程化方法,利用标准化模块,由简单到复杂构建具有期望功能的生物系统的方法。\n关键的工程化概念标准化建立生物功能的定义、建立识别生物部件的方法及标准生物部件的注册登记解耦将复杂问题分解成许多相对简单的、可以独立处理的问题,最终整合成具有特定功能的统一整体的过程抽提包括建立装置和模块的层次,允许不同层次间的分离和有限的信息交换、开发重设计的和简化的装置和模块、构建具有统一接口的部件库\n1.6合成生物学与相关生物学科与遗传工程的关系又称基因工程,在分子水平上对基因进行操作的复杂技术。是将外源基因通过体外重组后导入受体细胞内,使这个基因能在受体细胞内复制、转录、翻译表达的操作。与生物信息学的关系“信息爆炸—知识匮乏”合成生物学强调生物系统的设计和优化\n基因工程合成生物学将个别外源基因转移到某生物基因组内,使之能表达有益的蛋白质从头设计和构建自然界中不存在的人工生物体系较少使用数学工具在设计和构建人造生物体系时广泛使用各种数学工具来进行模拟,使得设计和构建的生物体系能正常工作在基因工程的实施过程中由于只转移个别外源基因,一般较少或不进行细胞网络分析改变了“转移一个基因,表达一种蛋白质”的模式,而通常是转移一组基因,因而要在更大规模更多层次上涉及到细胞网络,如代谢网络等\n与分子生物学和细胞生物学的关系分子生物学在分子水平上研究生命现象的科学。通过研究生物大分子(核酸、蛋白质)的结构、功能和生物合成等方面来阐明各种生命现象的本质。研究内容包括各种生命过程。比如光合作用、发育的分子机制、神经活动的机理、癌的发生等细胞生物学以细胞为研究对象,从细胞的整体水平、亚显微水平、分子水平等三个层次,以动态的观点,研究细胞和细胞器的结构和功能、细胞的生活史和各种生命活动规律的学科。合成生物学利用分子和细胞生物学的研究成果设计和重新构造新的生物分子、遗传线路乃至具有崭新功能的细胞,并在此基础上验证和检验分子和细胞生物学的理论。\n合成生物学研究举例“合成生物学”2004年被美国MIT出版的“TechnologyReview”评为将改变世界的10大新出现的技术之一(10EmergingTechnologiesThatWillChangeYourWorld)。短短几年的实践已经证明该预见的正确性。通过使用自然界提供的简单模块,许多合成生物学研究者在进行两方面的研究:建立有用装置;研究可以取代自然生物系统的人造生物系统。这些进展主要涉及如下几方面:(a)合成振荡器和开关;(b)人造细胞-细胞相互作用系统;(c)工程化的信号转导系统;(d)代谢途径工程;(e)通过蛋白质工程制造的新型生物传感器;(f)最小细胞及合成基因组。这方面的进展已经有许多专门的综述。\n(1)青蒿素的生物合成美国UCB化学工程系教授、劳伦斯国家实验室合成生物学中心主任Keasling本科曾获得化学与生物学双学位,后来又获得化学工程博士学位。所以在他从事代谢工程及合成生物学的研究中能很好地将化学、生物学、化学工程学相结合。在他从事抗疟疾药的生物合成研究中,始终把细胞当作微生物制药工厂(Microbialdrugfactories),进行设计、加工、集成、组装、控制。体现在合成生物学技术上包括DNA的合成、来自细菌、酵母及植物(青蒿Artemisiaannua)等多种基因及代谢途径的组装、多基因的精密调控等。他们关于抗疟疾药物生物合成的研究成果先后发表在2003年NatureBiotechnology和2006年的Nature上。合成生物学研究举例\n基因线路的应用举例什么是基因线路?一个典型的基因线路是基因双稳态线路,由两个蛋白质编码基因与两个相对应的启动子组成。线路如何设计?蛋白质1的表达抑制了蛋白质2的表达,系统只有蛋白质1存在;反之,蛋白质2的表达抑制了蛋白质1的表达,系统只有蛋白质2存在。可在双稳态线路中加入诱导物,促使系统在两个稳定状态之间任意翻转\n大肠杆菌照相术美国加州大学旧金山分校的合成生物学家C.Voigt设计基因线路当有红光照射时(相当于被摄物体的光亮部分),涂抹在琼脂基片上的菌苔形成的底片保持原色;当没有红光照射时,相当于被摄物体的黑暗部分,菌苔生成一种黑色沉淀物\n大肠杆菌计算机烧饼问题设想有一摞大小各不相同,烤面和背面任意取向的烤饼,欲把所有烤饼按从大到小的顺序排好,同时要求所有烤面朝上,规定每步只许改变一个或几个相邻烤饼的位置和方向,问最小要操作多少步才能完成任务6个烧饼-46080种排列组合12个烧饼-1.9万亿种排列组合\n生物学家的解决方案美国Davidson学院的K.Haynes用长短和方向(5’→3’或3’→5’)各不相同的DNA片断来代表烤饼。由重组酶Hin/hix来完成“烤饼”的换位和方向改变。设计一个基因线路,使当“烤饼”按要求排好后,抗四环素基因表达,反之则不表达。然后把这种工程化的大肠杆菌大量繁殖,使一个培养皿中达到数亿个菌体。每个细菌都是一部“计算机”,各自独立、互不干扰的进行“计算”。一段时间后用四环素处理菌体,没有完成排序任务的细菌都被杀死了,剩下的只有完成排序任务的菌体。从一开始到找到完成任务的菌体所需时间模拟了最少操作步数。\n大肠杆菌砷探测器在世界各地尤其是孟加拉国,饮用水中砷或亚砷酸盐往往超标,危害数百万人的身体健康。世界卫生组织(WHO)要求饮用水砷或亚砷酸盐的浓度不得超过10ppb。现有的砷检测方法难以达到这一精度。英国爱丁堡大学的合成生物学家设计了专用于检测砷的基因线路。在有低浓度砷或亚砷酸盐时(约5ppb),培养液呈中性;当有高浓度或亚砷酸盐时(约20ppb),培养液呈酸性,pH值可达5以下,可利用pH试纸或者pH指示剂检测。\n1.4生物系统的层次化结构生物系统的层次化结构是合成生物学工程化本质的又一典型体现。具有一定功能的DNA序列组成的最简单的BioBrick称为基因部件-Part。不同功能的基因部件按照一定的逻辑和物理连接组成复杂的生物装置-Device,不同功能的Device协同运作组成更加复杂的生物系统-System.含有多种不同功能System的生物体彼此通讯互相协调组成再复杂些的多细胞或细胞群体生物系统。\n1.4.1Part遗传系统中最简单、最基本的生物积块称为生物部件-Part。Part是指具有特定功能的核苷酸或者蛋白质序列,能够通过标准化组装方法与其他Part组装成具有更复杂功能的模块。Part按照其功能可以划分为终止子、蛋白质编码基因、报告基因、信号传递组件、引物组件、标签组件(tag)、蛋白质发生组件、转换器、启动子等类别。每一个Part都有一个标准的名字编码,我们可以很方便地从一块DNA元件的名字编码中判断出它在具体生物过程中所发挥的功能。\n常见的元件Part如下:启动子启动子(promoter,P)是操纵子(operon,O)的一个组成部分,专一地与RNA聚合酶结合并决定转录从何处起始的部位,控制基因表达(转录)的起始时间和表达的程度。启动子就像“开关”,与称为转录(transcription)因子的蛋白质结合,控制基因的活动。转录因子是“起始复合物(initiationcomplex)”的组成成分,指导RNA聚合酶的转录起始。生物中有许多启动子,如大肠杆菌约有2000个启动子,各启动子的效率并不相同,强启动子每2s启动一次转录,而弱启动子每10min才启动一次。细菌的启动子通常具有一些为RNA聚合酶与启动子相结合所必需的特定的结构保守区,其碱基变化会影响RNA聚合酶的识别能力和结合亲和力,控制转录水平的高低。原核表达系统中通常使用的可调控的启动子有Plac(乳糖启动子)、Ptrp(色氨酸启动子)、Ptac(乳糖和色氨酸的复合启动子)、T7噬菌体启动子等。真核生物的启动子与原核生物不同,而且启动转录的活性除需启动子外,还需某些外加序列。\nRBS-核糖体结合位点(ribosomebindingsite,RBS)是指mRNA分子中紧靠启动子下游、起始密码子AUG上游的一段非翻译区序列,用于结合核糖体以便开始转录。翻译起始密码位于它的下游位置,其功能是初始化翻译。原核生物的RBS中有SD序列,长度一般为4~9个核苷酸.富含G、A。该序列与核糖体16SrRNA的3’断互补配对,促使核糖体结合到mRNA上,有利于翻译的起始。RBS的结合强度取决于SD序列的结构及其与起始密码子AUG之间的距离,相距一般以4~10个核苷酸为佳,9个核苷酸为最优。由于核苷酸的变化能够改变mRNA5’端的二级结构,影响核糖体30S亚基与mRNA的结合自由能,从而造成蛋白质合成效率上的差异,因此SD序列的微小变化往往就会导致表达效率上百倍甚至上千倍的差异。SD序列(Shine-Dalgarnosequence):mRNA中用于结合原核生物核糖体的序列\n终止子是指结束基因转录的DNA信号序列。在一个基因的3’端或是一个操纵子的3’端往往有特定的核苷酸序列,具有终止转录的功能,这一序列称之为转录终止子,简称终止子(terminator,T)。所有原核生物的终止子在终止点之前都有一个回文结构,它转录出来的RNA可以形成一个茎环式的发夹结构。终止子按其作用是否需蛋白因子的协助至少可以分为两类:一类是不依赖ρ因子的终止子,这类终止子在序列上有一些共同的特点,即有一段富含GC的反向重复序列(invertedrepeatsequence),其后跟随一段富含AT的序列,因而转录生成的mRNA序列中能形成发夹式结构,后继一连串寡聚U序列。正是RNA聚合酶转录生成的这段mRNA的结构阻止RNA聚合酶继续沿DNA移动,并使聚合酶从DNA链上脱落下来,终止转录。另一类是依赖ρ因子的终止子,即其终止转录的作用需要ρ因子的协同,或至少是受ρ因子的影响,终止点前无寡聚U序列,回文对称区不富含GC。不同终止子的作用也有强弱之分。有的终止子几乎能完全停止转录;有的则只是部分终止转录,还有一部分RNA聚合酶能越过这类终止序列继续沿DNA移动并转录。如果一串结构基因群中间有这种弱终止子的存在,则前后转录产物的量会有所不同,这也是终止子调节基因群中不同基因表达产物比例的一种方式。转录终止的机制较为复杂,结论尚不统一。在构建表达载体时,为了稳定载体系统,防止克隆的外源基因表达干扰载体的稳定性,一般都在多克隆位点的下游插入一段很强的转录终止子。\n操纵子是细菌的基因表达调节装置,由启动子和其他DNA调节元件与串联的多个相关基因组成,由同一套调节蛋白调节。操纵子通常由2个以上的编码序列、启动序列、操纵序列以及其他调节序列在基因组中成簇串联组成。某些操纵序列是原核阻遏蛋白的结合位点,当操纵序列结合阻遏蛋白时会阻碍RNA聚合酶与启动序列的结合,或使RNA聚合酶不能沿DNA向前移动,阻遏转录,介导负调节(negativeregulation)。原核操纵子调节序列中还有一种特异DNA序列可结合激活蛋白,使转录激活,介导正调节(positiveregulation)\n1.4.2生物装置有了上述标准化的“Part”部件,就可以利用转录激活因子、转录阻遏蛋白、转录后机制(如DNA修饰酶)和Riboregulator(核糖核酸调节因子)等结合逻辑拓扑结构构建稍微复杂些的生物装置“Device”。生物装置通过调控信息流、代谢作用、生物合成功能以及与其他装置和环境进行交流等方式处理“输入”产生“输出”。\n利用iGEMRegistry提供的标准化系统量化方法,我们可以将一些生物装置进行标准化抽提,描述成如下形式:具有一定生物学功能,并且能够为外源物质所控制的一串DNA序列。报告基因(reporter)——其产物易于被检出的基因,在分子生物学实验中用于替换天然基因的位置,以检验其启动子及调节因子的结构组成和效率,常用的为各种荧光蛋白编码基因,如gfp(greenfluorecentprotein基因)等。转换器(inverter)——一种遗传装置,它在接收到某种信号时停止下游基因的转录,而未接收到信号时开启下游基因的转录。信号转导装置(signaling)——是指环境与细胞之间或者邻近的细胞与细胞之间接收信号和传递信号的装置。蛋白质生成装置(proteingenerator)——产生具有一定功能蛋白质的装置\n1.4.3生物系统为了得到更加复杂的调控行为或生物功能,可将装置以串联、反馈或者前馈等形式连接,组成更加复杂的级联线路或者调控网络,即所谓的生物系统(System)。自然生物系统中的调控级联线路是非常普遍的,如大肠杆菌和酵母菌调控网络中包含的级联线路。级联线路调控基因的递进式表达,可以触发大肠杆菌鞭毛、酵母菌孢子的形成或者控制细菌的细胞周期。在活体细胞中,许多信号转导和蛋白激酶通路也得益于级联过程来调控其活性。在果蝇和海胆等多细胞生物体中,许多时间顺序事件通常由级联过程来调控。\n级联线路具有许多非常重要的特点。例如,由蛋白质控制的级联线路对于渐变输入具有超敏感性的“全开或全关”响应,输入信号中非常小的变化即可以激励输出由低到高/由高到低的跳变在各种级联线路和调控网络中,最简单的形式是转录水平的调控系统。\n1.4.4细胞群体系统及多细胞系统由于基因表达过程中内源和外源噪声的影响以及其他细胞的作用,互不通讯的一组细胞即使起源相同也可能具有不同表型和异步行为,不可能互相协作,完成的生物功能也有限。为了更高效地实现人类期望的功能,需要多个细胞甚至多种细胞协同运作。利用细胞间通讯协调彼此的行为是目前工程化细胞群体系统的主要手段。相比于单细胞而言.细胞群体系统及多细胞系统的人工构建则复杂得多,不仅要考虑细胞间的协同,还要考虑信号分子的跨膜运输、环境因素的分布梯度等。\n群体感应此现象最早是在海洋中的一种发光弧菌(Vibriofischeri)中发现的。近年来的研究证明,微生物之间存在着信息交流,许多微生物都能合成并释放一种被称为自体诱导物(autoinducer,AI)的信号分子,胞外的AI浓度随微生物密度的增加而增加,当达到一个临界浓度时,AI能启动菌体中相关基因的表达,调控微生物的生物行为,如产生毒素、形成生物膜、产生抗生素、生成孢子、产生荧光等,以适应环境的变化,这一现象被称为群体感应现象。因这一现象只有在微生物密度达到一定阈值后才会发生,所以也称为依赖细胞密度控制的基因表达(celldensitydependentcontrolofgeneexpression)。\n在发光弧菌中由Plux启动子控制群体感应表达系统,而Plux启动子是由小分子信号控制的\n群体感应一般用于系统中群体细胞之间的同步响应,它提供了一种依赖细胞浓度调控基因表达的有效手段。合成生物学在此方面的初期尝试就是在大肠杆菌中开发细胞间的通讯模块以协调细胞群体行为。将费氏弧菌(Vibriofischeri)中响应群体感应的基因分别植入独立的发生细胞和接收细胞中。发生细胞表达Luxl催化AHL的合成。AHL信号分子自由扩散到接收细胞中和LuxR蛋白组成复合物激活目的基因转录,从而通过AHL信号分子的扩散实现细胞群体间的通讯和协作。\n第二部分合成生物系统的逻辑拓扑结构原核生物基因表达调控主要发生在转录水平,因此以转录水平的调控为主来介绍遗传线路的各种逻辑拓扑结构。转录调控的基本单元是操纵子,在转录水平上对基因表达进行调控取决于DNA的结构、RNA聚合酶的功能、蛋白因子及其他小分子配基的相互作用。对于这些拓扑结构的分类方式有很多种,大体可以归纳为串联、反馈和前馈等类型。其中,反馈可分为正反馈和负反馈;前馈可分为一致前馈和非一致前馈等。这些拓扑结构并不是生物系统所特有的,在电子线路、控制理论、计算机图论等领域中均有类似的拓扑结构,只是因为生物系统是由生物元件组成,具有不同于电子线路、控制回路等的特点。\n1.5串联串联是指上游模块的输出信号作为下游模块的输入信号的连接形式对于生物模块来讲,这种信号可以是小分子代谢物、蛋白质、mRNA等。\n1.6单输入主模块的输出作为下层分模块的输入。在单输入结构中,只有一个主模块作为下层模块的输入。\n这种结构在自然生物的转录基元(motif)中很常见,其主要功能是实现一组基因的共表达。此外还可以实现模块时序表达的功能,即模块1到模块N按预定义顺序表达:如果主模块代表激活因子,激活下层启动子模块,启动子序列和结合位点不同,导致各个启动子激活阈值不同,当主模块的活性缓慢递增时,阈值最低的启动子先启动,依此类推,阈值最高的启动子最后启动,实现基因表达的时序控制。同理,当主模块代表阻遏子基因时,可以实现类似的时序控制。通过高时间分辨率的检测手段可以观测到在大肠杆菌中有许多系统具有SIM结构。这一结构具有一个很重要的属性——时间序列与基因功能序列吻合,即在一个多基因过程中,越早需要的蛋白质/代谢物,它的基因越先被激活。这种时序控制可以防止蛋白质产物在需要之前过早生成,实现能源的节约,避免不必要的交叉干扰。\n在大肠杆菌的精氨酸生物合成系统中,阻遏蛋白ArgR调控多个编码精氨酸生物合成过程中所需要的酶的操纵子精氨酸合成的主要途径是利用谷氨酸盐作为前体的,此外还需要谷氨酰胺和天冬氨酸,其产物通过另外两条无支路的途径合成,其并最终会合于主路径。\n1.7多输入多输入模块(multiinputmodule,MIM)结构也称为“密集交盖调节网”(denseoverlappingregulons,DOR)。以转录水平的调控为例,这种结构表示一组调控因子共同控制一组基因。大肠杆菌中有很多DOR结构,负责碳的利用、厌氧环境下生长、胁迫响应等广泛的生物功能。类似的结构在酵母菌中也有很多。\n由图中可以看出,MIM可以看作是一个逻辑门阵列,多路输入进行组合运算以后控制下游模块。因此,为了充分理解模块之间的调控机制,仅用箭头来表示信号的传递是不够的\n在8个子图中,只有子图C、G中两个调控因子对被调控基因的作用是完全一致的\n1.8反馈反馈是控制理论中最重要的概念之一。从抽象的信号层面讲,反馈是指系统的输出会反过来影响系统的输入,进而影响自身的一种控制机制。这种输出对于输入的影响可能会导致最终输出的降低,即所谓的负反馈;也可能会导致最终输出的增加,即所谓的正反馈。反馈的正负特性由具体的网络动力学特性决定。由于从概念层面上讲,信号的传递形成了一个闭合的回路,因此也常称此结构为“反馈回路”。“反馈”在现实社会中的应用极其广泛,大到各种政策的调控、大规模化工厂的流程控制、各种生物代谢网络,小到基因转录的调控都有反馈结构存在。\n在最早的遗传线路研究中,人们发现基因编码的蛋白质可能会反过来增加或者阻遏上游基因的开启。因此,人们借鉴控制领域中的相应理论和概念来描述这种现象,称其为“反馈”。\n遗传线路中也存在着两种基本反馈方式,即负反馈和正反馈。M.A.Savageau经过研究,较早提出了“负反馈的调控比开环或者正反馈的调控要更稳定”的论断后来A.Becskei等在实验中证明了这两种基因反馈的存在;A.Becskei等同时还证明了正反馈对于生物系统的双稳态行为具有重要作用正反馈:基因表达的产物会反过来促进相关基因表达。负反馈:基因表达的产物会反过来抑制相关基因表达。\n\n如果基因的表达量随着信号分子的加入而增加,这种遗传线路就被定义为可诱导的;如果基因的表达量随着信号分子的加入而减少,这种遗传线路就被定义为可阻遏的。与分解代谢有关的遗传线路倾向于被代谢中间产物所诱导,即代谢中间产物对于分解代谢具有正反馈作用。与之相反,合成遗传线路倾向于被一种代谢途径的终产物抑制,即终产物通过负反馈抑制自身合成,否则将会造成合成酶的过度浪费。反馈的引入增加了基因线路调控的精确性,线路的稳态输出不再是输入的简单函数\n1.9前馈前馈控制也称预先控制或提前控制,其基本原理就是测取进入过程的扰动量(包括外界扰动和设定值变化)。并按照其信号产生合适的控制作用去改变控制量,使被控制量维持在设定值上。前馈控制与反馈控制的最大区别就在于,它是在偏差出现之前就采取控制措施,而后者则是在偏差出现之后才采取纠正措施。前馈结构的特点是干扰信号进入系统后分成干扰通路和补偿通路两条不同途径影响最终变量\n基因线路中利用“前馈”来表示上游基因通过两条不同途径影响下游基因的控制结构。根据这两条通路对最终基因的影响效果是否一致,前馈结构还可进一步划分为“一致”前馈(coherent)和“非一致”前馈(incoherent)两类。“一致前馈”(coherentfeedforward)模块中直接和间接调控途径对输出模块的作用相同;“非一致前馈”模块(incoherentfeedforwardmodules)中直接和间接调控对输出模块的作用相反\n最简单的前馈转录基元(motif)由三个基因组成:基因X调控基因Y和Z,基因Z同时受到基因X和Y的调控X、Y、Z基因之间彼此的促进和抑制关系不同,因此三个元素组成的前馈具有8种可能的结构类型A所示为4种“一致前馈”类型,即X对Z的直接调控促进X通过Y对Z的间接调控,两条支路对Z的调控效果是一致的。B所示为4种“非一致前馈”类型,即X对Z的直接调控抑制X通过Y对Z的间接调控,两条支路对Z的调控效果是不一致的\n理解前馈结构必须很好地理解基因X和Y对Z的共同作用。可以用两种逻辑门来模拟X和Y对Z的调控作用:“与”门表示Z的激活需要X和Y同时满足要求,如图C所示;“或”门表示X和Y中只要有一个满足要求即可激活Z,如图D所示。\n其他拓扑结构\n常用性能指标稳定性(stability)反映在暂态扰动消失后的时间响应性质上,对于稳定性的分析,有助于人们了解系统能够承受的最大干扰的幅值和频率,从而对实验设计有一定的指导意义。定义为:系统在任何足够小的初始偏差的作用下,对于暂态扰动的时间响应随着时间的推移而逐渐衰减并趋向于零,则该系统是稳定的;否则,该系统是不稳定的。稳定性是相对而言的,再稳定的系统所能承受的外界干扰也是有限的。如果外界干扰幅值过大,系统会因无法承受而崩溃,或者发生结构改变。\n鲁棒性(robustness)就是系统的健壮性。它是在异常和危险情况下系统生存的关键。所谓“鲁棒性”,是指系统在一定(结构、大小)的参数摄动下,维持某些性能的特性。例如,计算机软件在输入错误、磁盘故障、网络过载或有意攻击情况下,能否不死机、不崩溃,就是该软件的鲁棒性。鲁棒性和稳定性是从两个不同角度衡量合成系统抵御宿主细胞内外环境扰动的能力。鲁棒性侧重扰动导致模块自身结构参数变化时维持原来状态的能力;而稳定性则侧重模块受到外界扰动后维持稳定的能力,模块也许会从原来的稳定状态过渡到新的稳定状态,但仍然维持稳定的能力\n响应快速性所谓的响应快速性就是处于稳定状态的系统对于外部环境改变的快速反应能力,也是系统在受到扰动后迅速进入稳态的能力。对于生物模块来讲,任何输入信号,无论是信号分子、蛋白质,还是代谢物都会降解、排出或者转化成其他物质。生物模块必须要在这些输入信号消失之前做出响应,才能确保自身功能的发挥响应快速性可以通过改变具体的基因序列或者生物大分子构象等多种方法来改进。而且,响应的快速性是建立在系统的鲁棒性和稳定性基础上的,只有保证模块或者系统不被分解、不发散,快速性才有意义。因此,对于生物模块的性能分析还足以稳定性和鲁棒性为主\n1.5稳定性分析目前对于系统的稳定性分析还是主要建立在系统模型的基础之上,对于模型的精度具有一定的依赖性。一般可以采用两种方法来分析其稳定性:解析法和数值法。解析法分析通常需要繁琐的运算,对于没有解析解的模型来说甚至根本行不通。因此通常采用数值解法来分析系统的稳定性。相位面分析是数值分析的常用方法\n1.6鲁棒性分析/敏感性分析鲁棒性分析关心的是系统参数发生摄动时,系统能否保持原有的性能和状态,因此需要分析参数变动对于状态的影响,即参数敏感性分析。敏感性分析并不能得出真实的速率常数,只是辅助确定哪个参数更重要。在知识不完备情况下进行系统设计时,确定对系统输出具有最显著影响的参数是至关重要的:如果需要维持系统性能。则应尽量维持最敏感的参数不变;如果需要改变系统性能,则改变最敏感参数可能是最便捷的途径,因为系统输出通常只对少数几个参数敏感,相应突变的候选反应数量则完全可能缩小到实验可以处理的范围内\n1.7提高系统稳定性和鲁棒性的机制生物系统各种各样的内源和外源噪声都能影响合成生物模块在活体内发挥期望的能。基因表达中内源因素引起的噪声主要是指单个基因水平上生化反应的随机性;而外源噪声则包括代谢物、核糖体以及聚合酶等细胞组分的波动。有些研究表明,基因表达内源噪声引起的干扰一般要快于细胞周期,而外源噪声引起的干扰则具有较长的时间尺度且对合成模块的功能具有更显著的影响。噪声对于具体合成模块的影响不仅取决于噪声本身的属性。也取决于模块的属性。在进行装置及模块功能模拟和仿真时考虑噪声的影响对于生物系统设计按照预期功能工作非常重要。可以利用某些机制来提高系统的稳定性和鲁棒性。\n冗余生物系统确保预期行为的最简单方法即是冗余性。这方面的例子有很多,例如,胰岛紊信号转导通路和酵母菌大部分代谢途径都具有一定的冗余性。当用在连通性能完好的节点上时,冗余性能提高无标度网络的稳定性。细胞的代谢系统大多数属于无标度网络,因此这一机制非常有用\n反馈控制许多网络都具有一些防止系统输出达到无法控制情况的机制。正如单回路控制系统比开环控制系统具有更强的鲁棒性一样,转录级联线路比单基因转录系统更能抵抗外源噪声的影响。某些情况下,反馈能够增加系统的超敏感性,而生物模块因超敏感性而具有的近似离散的响应,使得构建的复杂系统具有一定的离散性和鲁棒性。负反馈回路能够抑制噪声干扰并提高系统的稳定性。正反馈通常具有较差的稳定性,但由于能够提高系统的敏感性,因此对于系统的鲁棒性也有至关重要的影响。一个非常微弱的信号能够通过正反馈环放大并启动期望行为,导致快速或者局部化的切换/开关。正反馈系统不能改善稳定特性,但能够造成系统“爆炸”。因此,当不再需要正反馈的行为时,必须有机制能够及对断开正反馈回路。当将转录速率限制在特定水平时,正反馈能够有效消除抑制转录速率的扰动,使其稳定在有限水平\n特异性和保真度信号通路的特异性和保真度能够帮助我们衡量系统各组分间相互作用的程度。\n\n\n\n\n1.2基因调控开关除了逻辑门遗传线路外,最基本的基因表达调控部件即是控制基因表达的各种“基因开关”。基因开关是指某种化学诱导物存在或缺乏时,或者在两个独立的外源刺激作用下,基因处于两种可能状态中的一种的系统。\n1.2.1转换开关转换开关(inverter)因其输出是输入的转换函数而得名,即输入为低时输出是高,反之亦然。天然系统中有许多基因调控的转换开关存在,如正控阻遏系统(即效应物分子的存在使激活蛋白处于非活性状态,转录不进行)即可以看做是天然的转换开关:效应分子输入为高时,基因不表达,系统输出为低;效应分子输入为低时,基因表达,系统输出为高\n1.2.2双相开关噬菌体中通过一个双向操纵子OR来维持溶源状态,这个操纵子由阻遏蛋白CI和反阻遏蛋白Cro(分别由cI基因和cro基因编码)结合到相邻的三个结合位点(OR1,OR2,OR3)来调控CI蛋白对cro基因的转录起着负调控作用,对cI基因本身的转录则既有正调控作用又有负调控作用,即我们所说的双相开关(biphasicswitch)\n双相开关(biphasicswitch)机理\n1.2.3Riboswitch生物体能够通过各种精巧的机制控制基因表达的时间和数量,几年前发现的Riboswitch就是这些精巧机制中的一种。Riboswitch主要是通过核糖核酸(RNA)构象改变来实现“开关”的功能,阻止或开启目的蛋白的生成大部分Riboswitch只有一个识别靶向配体的结合位点或适配体(aptamer);适配体一般位于基因表达区域附近,当其与代谢产物结合时,会改变自身结构,在转录或翻译水平上行使基因调控的功能Riboswitch可被一种调节sRNA(此处指小片段的调节RNA,非编码RNA的一种,对于真核和原核生物的调节作用非常重要)“打开”,重新激活转录或翻译\nRiboswitch在进化上可能是RNA世界遗留的分子化石,广泛存在于革兰氏阴性菌和革兰氏阳性菌的代谢相关基因中,在真菌和植物中也有发现。Riboswitch调节基因表达不需要任何蛋白因子作为中介,主要参与调节维生素、氨基酸和核苷酸等基础物质的代谢过程,因此可用于研究基因功能、开发新型药物及进行基因治疗\nT.S.Bayer和C.D.Smolke等设计的短链RNA基因调控系统\n1.2.4双稳态开关双稳态开关也称“拨动开关”,可通过人为调控实现基因线路在两种不同稳定状态间的切换经典的转录水平双稳态开关由两个启动子组成,每个启动子的“开”和“关”状态之间具有明显的界限,只有通过瞬态的诱导因素变化才能切换基因拨动开关至其中的一个稳态,且在移除其输入激励后仍会维持原来的状态。双稳态开关对于输入信号在一个较宽范围内的变化不敏感,因此对信号扰动具有一定的鲁棒性。通常由一种状态切换到另一种状态时,需要时间来进行转换,因此双稳态开关起始状态为“开”或“关”时会得到不同的响应曲线。这一特性使双稳态开关具有“记忆”的功能,可能会导致细胞状态的不可逆转性。\nJ.J.Collins课题组构建的经典的双稳态开关基因线路设计图中有两种不同的启动子——启动子1和启动子2,它们分别控制着阻遏子2和阻遏子1基因序列的表达状态;而阻遏子2和阻遏子1基因表达的产物又会分别抑制启动子2和启动子1的启动。阻遏子1下游报告基因的表达产物可以作为输出信号表征系统目前所处的状态。由于上下游基因的启动和相互抑制,整个系统共有下面两种稳定的输出状态。\n系统的两个外部输入信号——诱导物1和诱导物2,分别能解除阻遏子1基因的表达产物对启动子1的抑制作用和阻遏子2基因的表达产物对启动子2的抑制作用。他们的加入可以实现系统在两种状态之间的转换,是人工实现状态切换的手段当系统稳定于状态A时,通过加入诱导物2,可以解除阻遏子2对启动子2的抑制作用,使得阻遏子1表达,从而抑制启动子1,终止阻遏子2的表达。一段时间后,系统从状态A转换至状态B,而即使解除诱导物2的输入,系统依然会稳定与状态B同理,加入诱导物1作用一段时间,也可使系统从状态B切换到状态A。如果将诱导物1和2的加入看作是对开关的“拨动”,那么每次拨动后,系统都会稳定于相应的状态,而不需要其他外界信号来维持\n基因线路实现\n基因线路实现研究者利用不同的载体,分别构建了两类不同的双稳态开关。一类是利用方案A和方案B在pTAK载体上构建双稳态开关,称为pTAK类一类是利用方案A和方案C在pIKE载体上构建双稳态开关,称为pIKE类。无荧光蛋白有荧光蛋白IPTG温度无荧光蛋白有荧光蛋白IPTGaTc\n功能的检测所有的双稳态开关都被转入大肠杆菌JM2.300菌种中验证其功能。带有双稳态开关质粒的大肠杆菌在培养基中培养23.5h,分别用不同的诱导物诱导一段时间后再转入新的培养基中培养。通过检测不同时刻系统中荧光的量来检测双稳态开关的功能。\n\n1.3Repressilator基因振荡是一种基因调控机制,由振荡的幅度和周期来决定基因表达的时间。这种基因表达的时间控制可以实现在大规模基因网络中仅利用少量的调控因子即可调控相对较多的基因,从而实现对复杂细胞行为的调控。在转录水平上,如果将三个表达产物相互抑制的基因模块串联成一个环状结构,利用基因模块间的彼此抑制和解抑制即可实现振荡器的功能。这就是M.B.Elowitz等构建的基因振荡器。该Repressilator实现了在没有外加输入作用下,生物系统中基因表达按照特定的周期变化的功能\n遗传线路设计M.B.Elowitz等设计的遗传线路如下图。采用了三个启动子,每个启动子所控制的基因均能抑制下游最近邻一个启动子的启动\n第二部分其他方面的合成生物学基础研究控制群体数量的基因线路在大肠杆菌中构造了利用信号感应系统调控细胞凋亡来控制细胞群体浓度的基因线路。将单个细胞基因线路的作用结果扩散至细胞群体中,通过以群体行为的统计学结果作为基因线路的表征,实现对细胞群体浓度的人工调控。细胞凋亡又称程序性细胞死亡,是一种高度调节的生理性细胞死亡过程。与坏死不同,细胞凋亡是指细胞接受某种信号或受到某些因素的刺激后,会发生主动的、由基因调控的消亡过程。由于细胞凋亡过程通常伴随有重要的生物现象。\n